MI-07374
MI-073741999
![]() MI-07374
MI-07374
1999
Michael J. McDonough,
Graduate Student Department of Horticulture
Daniel Gerace,
Research Fellow Department of Entomology
Mark E. Ascerno,
Extension Entomologist and Department Head Department of Entomology
![]()
|
Western flower thrips (WFT) are probably the most serious pest of floriculture crops in the world (Parrella 1995b). WFT damage plants directly by feeding and laying eggs on the plant, and indirectly by acting as vectors for tospoviruses such as tomato spotted wilt virus (TSWV) and impatiens necrotic spot virus (INSV). A recent survey of Midwest greenhouse operators identified WFT as the most difficult greenhouse pest to manage (Wawrzynski and Ascerno, in review). Daughtrey (1997) summarized the WFT-tospovirus problem at a recent conference of the Society of American Florists as follows: “growers who recognize the disease symptoms and concentrate on disease management still may suffer some losses from the virus periodically, but those who fail to become well informed about thrips and virus management are courting disaster.”
AppearanceAdult WFT are less than 1/20th of an inch (<2mm) long (Govena and Flint 1996). Male adult WFT are light yellow, and have narrow abdomens; female adult WFT are larger than the males and vary in color from light yellow, yellow with brown splotches, to dark brown (Parrella 1996). Adult WFT have slender bodies with two sets of narrow, clear, nearly veinless wings that have dark, hairy, fringes (Johnson and Lyon 1991). WFT is the most prevalent species of thrips in U.S. greenhouses.
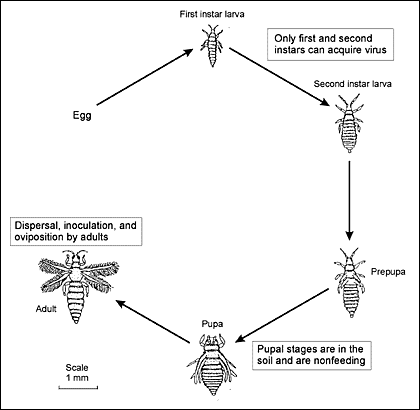
Biology, Life Cycle, and BehaviorDepending on environmental conditions and nutrient levels, female WFT lay 130–230 eggs during their lifetime (Robb 1989). The female WFT has an external ovipositor with two opposable serrated blades that are used to cut through the plant epidermis and deposit eggs in the tissues below (Childers and Achor 1995). Eggs are deposited in leaves, bracts, and petals and hatch in 2 to 4 days (Pfleger et al. 1995). The eggs of WFT are fairly well protected and few pesticide sprays are effective against them. WFT do not overwinter outdoors in the Upper Midwest but overwinter in soil and clover in the Mid-Atlantic states and southern Pennsylvania (Felland et al. 1995). Under greenhouse conditions, WFT probably reproduce continuously throughout the year. WFT give birth to large numbers of young, have rapid reproductive cycles, and increase their population faster than their predators, so early detection and control is essential (Mound and Teulon 1995). The life cycle of WFT consists of five stages: egg, two feeding larval instars, nonfeeding prepupa and pupa instars, and adult (Figure 1). The development time from egg to adult is 7 to 13 days when temperatures range from 62 to 98F (Robb 1989).
Plant DamageLeaf scars and specks of black feces are a good way of diagnosing the presence of WFT on plants. Depending on the host species, feeding injuries occur on fruit, flowers, flower buds, leaves, and leaf buds (Childers and Achor 1995). In some host species, WFT feeding causes flower or leaf buds to abort or emerging leaves to become distorted. Feeding does not always result in immediately visible damage because many flowering species do not show injury until flower buds open. When WFT feed on flowers, symptoms include streaks and discoloration of the petals—with dark flowers showing light streaks and light flowers showing dark streaks (Pfleger et al. 1995).
Mouthpart Anatomy and Plant Virus TransmissionThe mouthparts of WFT have been variously described as “rasping-sucking” or “piercing-sucking.” Recent research, however, shows WFT have piercing-sucking, multi-purpose mouthparts. The mouthparts are used to pierce leaves, flowers, seeds, pollen grains, and fruit, as well as to drink open liquids such as nectar, water, or insect secretions (Kirk 1995). Moreover, WFT are opportunistic feeders and eat mite eggs, adult mites, other thrips species, and even members of their own species. The anatomy of the mouthparts explains why WFT carry tospoviruses from plant to plant. In WFT the mouthparts are composed of a single mandibular stylet, which extends to punch feeding holes in the plant, and two maxillary stylets. These stylets, which are extended or retracted independently or simultaneously of the mandibular stylet contain a feeding/salivary channel (Mound and Teulon 1995). During feeding, saliva is injected into the plant cell and the contents withdrawn (Childers and Achor 1995). When tospoviruses are present in the saliva, these are transmitted to the plant during the feeding process.
TospovirusesTospoviruses are a major problem in commercial greenhouses for a number of reasons. Tospoviruses infect a very wide range of plants, cause significant economic losses when infections occur, and are transmitted from plant to plant by WFT—a common pest of commercial greenhouses. There are two major types of tospovirus, the tomato spotted wilt virus (TSWV) and the impatiens necrotic spot virus (INSV), which can be identified using on-site test kits. The best way to control tospoviruses in the greenhouse is to eliminate WFT. Host Plant Species and Virus Transmission Seven species of thrips are vectors for TSWV, but the only confirmed vector for INSV is WFT (Daughtrey et al. 1997). WFT acquire tospoviruses by feeding on infected host plants during the two larval instars and remain infected for life (Bandla et al. 1994). Adult WFT, however, are not infected by feeding on tospovirus-infected plants because tospoviruses pass right through the gut (Bandla et al. 1998; Ullman 1996). As a result, tospoviruses do not enter the salivary fluid and, thus, cannot infect plants (German et al. 1992). A partial list of plant species infected by tospoviruses is shown in Table 1 on the back of the accompanying laminated card that is part of this folder. (The laminated card can be bought separately as publication # MI-7375-C.) Host plants are categorized by crop type such as bedding, foliage, non-ornamental, ornamental, and perennial plants. A “+” notation in the respective TSWV or INSV column indicates the plant is susceptible to that particular virus. Some plants, such as gloxinia, are very sensitive to INSV. Gloxinia seedlings are rapidly killed and, when older plants are infected, total crop losses are not unusual (Daughtrey et al. 1997). Several host plants are susceptible to both TSWV and INSV, but it is important to note that not all plants susceptible to TSWV or INSV are hosts for WFT. Although WFT may probe non-host plants, they do not continue to feed and do not lay eggs. If thrips do not lay eggs (even though that plant is susceptible to tospovirus), that particular plant is not considered as a reservoir for either tospovirus. A complete host list for WFT has not been established, but finding any WFT larvae during plant inspections serves as warning that control measures are needed. Tospovirus Symptoms in Plants The symptoms of tospovirus infections in floral crops are listed below (Daughtrey and Casey 1998). The presence of one or more of these symptoms alerts you to potential tospovirus infections.
Tospovirus infections may be systemic (i.e., virus symptoms are spread throughout the plant) or non-systemic (i.e., the virus symptoms are confined to a specific part of the plant). Tospoviruses, however, may be present even though the plant shows no symptoms. Using On-Site Test Kits to Detect Tospoviruses Currently, the only kit available for on-site testing is the Agdia QTA-Tospo test kit, manufactured by Agdia Incorporated, Elkhart, Indiana. This kit uses a linked immuno-sorbent assay (ELISA) to detect the presence of specific viral proteins in the sample tissue. Separate kits are available from Agdia to detect TSWV and INSV. Another option for detecting tospoviruses is to send a plant sample to a plant disease diagnostic clinic. Samples may be sent to the University of Minnesota Plant Disease Clinic, 495 Borlaug Hall, 1991 Upper Buford Circle, St. Paul, MN 55108 ($35 for the first sample, $5 for additional samples). However, a detailed study of tissue samples sent for analysis to a testing laboratory in Long Island suggests that only 5 to 15 percent of the samples actually contain viruses (Daughtrey 1997), and another problem with testing for viruses is that some host plant species do not express TSWV or INSV symptoms until long after the initial infection. Cyclamen, for example, can show no symptoms for up to two months after infection (Allen and Matteoni 1988, cited in Daughtery 1996). Distribution and Treatment of Tospoviruses in the U.S.
Managing WFT and TospovirusesWFT is usually controlled in commercial greenhouses using a combination of sanitation (preventing WFT from entering greenhouses in the first place), cultural controls (managing crop plants, crop plant wastes, weeds, soil, and environmental conditions to eliminate WFT), chemical controls, and biological control (using other organisms, such as insects and fungi) to eliminate WFT. These methods, in combination with early warnings about WFT and tospovirus outbreaks using indicator plants, provide excellent control of both pests. Managing WFT is difficult because WFT
Physical Exclusion
Exclusionary screening enabled the Paul Ecke Poinsettia Ranch to reduce pesticide applications by 40 percent and still maintain a high level of control in poinsettia stock and New Guinea impatiens production areas (Hall 1995). Cultural Controls
Some growers apply a layer of hydrated lime thick enough to cover the soil beneath the benches because hydrated lime reduces algae, weeds, and insects in this critical area. Although dry hydrated lime may be applied, it may be easier to spray a slurry composed of 1 to 2 pounds per gallon of water (Lindquist 1998). As an alternative to soil sterilization, close greenhouse vents, keep the house fallow for seven days between crops to accelerate the development of WFT pupae in the soil, and eliminate the emerging adults (Parrella 1995b). Heating the greenhouses to temperatures in excess of 102F for two days will kill adult thrips (Parrella 1995b). Good Sanitation Controls Tospoviruses
The worldwide spread of WFT and TSWV/INSV is caused in two ways: by shipping tospovirus-infected plants (both propagative and mature) and by shipping plant materials infested with tospovirus-carrying WFT (Parrella 1995a). To minimize the chance of infection, segregate incoming plant material and quarantine plants until they can be inspected. Choose plant vendors with care, and only buy from vendors who have established virus prevention programs. Monitoring and Indicator Plants Petunia plants (Petunia x hybrida) are excellent early indicators for the presence of WFT feeding and the transmission of tospoviruses because petunias are not systemically infected with either TSWV or INSV (Ullman 1998). In response to a tospovirus infection, petunias show a hypersensitive response—a rapid death of plant tissues that also kills the invading virus (Robb et al. 1998). The following petunia cultivars are excellent indicator plants for the detection of tospoviruses (Robb et al. 1998; Daughtrey et al. 1995).
To set up a monitoring program using petunias, remove flowers from the indicator plants before placing them in production greenhouses because petunia flower petals do not express local lesions and attract WFT away from the leaves (Robb et al. 1998). Flag the indicator plants with blue pie pans or metal sheets to increase the effectiveness of the indicator plants since WFT are more sensitive to blue than to other colors. Look for WFT feeding scars, which are whitish and have an irregular outline. Brown or black-edged lesions will develop on the edges of thrips feeding scars within 3 days if a tospovirus has been transmitted (Ullman 1998). The accompanying laminated card (which is part of this folder, but can be bought separately as publication #MI-7375-C) shows photographs of uninfected feeding scars and feeding scars that have developed into lesions because tospoviruses are present. If a tospovirus outbreak occurs in the greenhouse, look for patterns of injury that correlate with variations in air movement, humidity, and temperature (Daughtrey 1998). Control measures include removal of infected plants, direct WFT control measures such as spraying, or exclusion of thrips. “Infected plants should be placed in bags or other containers at the bench to avoid spreading viruliferous [virus-containing] thrips to other areas of the greenhouse during the process of removal” (Moyer and Daub 1994). Biological Controls
Use of biological control methods should only be implemented as part of a well-planned IPM program. Biological control methods, however, should not be used when TSWV or INSV is already present in the greenhouse. In particular, when INSV is a recurring problem, biological control is inappropriate (Matteoni 1995). Chemical Control Controlling WFT with Pesticides When chemical control is necessary, spray pesticides 2 to 3 times over a period of five days (Robb and Parrella 1995; Daughtrey et al. 1995). Please note, this recommendation assumes greenhouse temperatures range from 21–29.5C (70–85F). When two sprays are used, reduce the spray interval to 3 days if greenhouse temperatures are higher, and increase the spray interval to 7–10 days when greenhouse temperatures are lower (Robb and Parrella 1995). After spraying, continue to monitor WFT population levels to determine if continued pesticide applications are necessary. Eggs and pupal stages are unaffected by pesticide sprays, so make sure the spraying program lasts long enough to include emerging larva and adults. Smaller droplet sizes are recommended because WFT prefer to feed on flowers and flower buds, and smaller droplets are more effective at penetrating flower parts (Robb 1995). Sugar is often used as a feeding stimulant in tank mixes but there is no scientific data to support the practice. Preventing Resistance
Pesticides labeled for greenhouse use against WFT are listed in Table 1. Please note, the table shows the class of each pesticide to help you plan your pesticide rotation schedule. Note carbamates and organophosphates have the same mode of action and should be treated as the same class. Tank mixes increase resistance problems and should be avoided.
ReferencesAllen, W. R., and J. A. Matteoni. 1988. Cyclamen ringspot: epidemics in Ontario greenhouses caused by tomato spotted wilt virus. Can. J. Plant Path. 10: 41-46. Bandla, M. D., D. M. Wescot, D. E. Ullman, T. L. German, and J. L. Sherwood. 1994. Use of monoclonal antibody to the nonstructural protein encoded by the small RNA of the tomato spotted wilt virus to identify viruliferous thrips. Phytopathology 84: 1427-1431. Bandla, M. D., D. M. Wescot, D. E. Ullman, T. L. German, and J. L. Sherwood. 1998. Interaction of tomato spotted wilt virus (TSVW) glycoproteins with a thrips midgut protein, a potential cellular receptor to TSWV. Phytopathology 88: 98-104. Childers, C. C., and D. S. Achor. 1995. Thrips feeding and oviposition injuries to economic plants, subsequent damage, and host responses to infestation. In B. L. Parker, M. Skinner and T. Lewis [eds.], Thrips Biology and Management. Plenum, New York, pp. 31-51. Daughtrey, M. L. 1996. Detection and identification of tospoviruses in greenhouses. Acta Horticulture 431: 90-98. Daughtrey, M. L. 1997. Real-world disease management for fungi and viruses: some serious current problems caused by cercospora, alternaria, phytophthora, and INSV. Proceedings for the Thirteenth Conference on Insect and Disease Management on Ornamentals, 22-24 February, 1997, Orlando, Florida. Society of American Florists, Alexandria, VA, pp. 43-52. Daughtrey, M. L. 1998. Don’t make mistakes with tospoviruses! Grower Talks 61(13): 104. Daughtrey, M. L., and C. Casey. 1998. Highlights from SAF’s pest conference. Grower Talks 61(14): 44-46. Daughtrey, M. L., R. K. Jones, J. W. Moyer, M. E. Daub, and J. R. Baker. 1997. Tospoviruses strike the greenhouse industry: INSV has become a major pathogen on flower crops. Plant Disease 81: 1220-1230. Daughtrey, M. L., R. L. Wick, and J. L. Peterson. 1995. Compendium of Flowering Plant Diseases. APS Press, St. Paul, MN. Felland, C. M., D. A. J. Teulon, and L. A. Hull. 1995. Overwintering and distribution of western flower thrips in the Mid-Atlantic United States. In B. L. Parker, M. Skinner and T. Lewis [eds.], Thrips Biology and Management. Plenum, New York, pp. 461-464. German, T. L., D. E. Ullman, and J. W. Moyer. 1992. Tospoviruses: diagnosis, molecular biology, and vector relationships. Annu. Rev. Phytopath. 30: 315-348. Govena, P., and M. L. Flint 1996. Thrips: home and landscape (http://www.ipm.ucdavis.edu/PMG/PESTNOTES/pn7429.html) (accessed March 5, 1999) Hall, J. 1995. Putting pest management to work. Proceedings for the Eleventh Conference on Insect and Disease Management on Ornamentals, 18-20 February, 1995, Fort Meyers, FL. Society of American Florists, Alexandria,VA, pp. 59-61. Johnson, T. J., and H. H. Lyon. 1991. Insects that Feed on Trees and Shrubs. Cornell University Press, Ithaca, NY. Kirk, W. D. J. 1995. Feeding behavior and nutritional requirements. In B. L. Parker, M. Skinner, and T. Lewis [eds.], Thrips Biology and Management. Plenum, New York, pp. 21-29. Law, M. D., and J. W. Moyer. 1990. A tomato spotted wilt-like virus with a serologically distinct N protein. J. Gen. Virology 73: 2125-2128. Lindquist, R. K. 1998. Fungus gnats and shore fly management. Proceedings for the Fourteenth Conference on Insect and Disease Management on Ornamentals, 21-23 February, 1998, Del Mar, CA. Society of American Florists, Alexandria, VA, pp. 45-51. Matteoni, J. A. 1995. Impatiens necrotic spot virus: update and integrated control. Proceedings for the Eleventh Conference on Insect and Disease Management on Ornamentals, 18-20 February, 1995, Fort Meyers, FL. Society of American Florists, Alexandria, VA, pp. 1-5. Mound, L. A., and D. A. J. Teulon. 1995. Thysanoptera as phytophagous opportunist. In B. L. Parker, M. Skinner and T. Lewis [eds.], Thrips Biology and Management. Plenum, New York, pp. 3-19. Moyer, J. W., and M. E. Daub. 1994. Tomato spotted wilt virus/impatiens necrotic spot virus: where we’ve been and where we’re going. Proceedings for the Tenth Conference on Insect and Disease Management on Ornamentals, 19-21 February, 1994, Dallas, TX. Society of American Florists, Alexandria, VA, pp. 86-93. Parrella, M. P. 1995a. IPM-approaches and prospects. In B. L. Parker, M. Skinner and T. Lewis [eds.], Thrips Biology and Management. Plenum, New York, pp. 357-363. Parrella, M. P. 1995b. Thrips management guide part I: prevention and control. Grower Talks 58: 30-38. Parrella, M. P. 1996. Introduction: western flower thrips—identification, biology and research on the development of control strategies. Proceedings for the Twelfth Conference on Insect Management on Ornamentals, 17-19 February, 1996, San Francisco, CA. Society of American Florists, Alexandria, VA, pp. 17-28. Pfleger, F. L., M. E. Ascerno, and R. Wawrzynski. 1995. Tomato spotted wilt virus. Minnesota Commercial Flower Growers Bulletin 44(2): 1-7. Powell, C. C., and R. K. Lindquist. 1997. Ball Pest and Disease Manual: Disease, Insect, and Mite Control on Flower and Foliage Crops, second edition. Ball Publishing, Batavia, IL. Robb, K. L. 1989. Analysis of Frankliniella occidentallis (Pergande) as a pest of floriculture crops in California greenhouses. Ph.D. dissertation, University of California, Riverside, Riverside, CA. 135 pp. Robb, K. L. 1995. Controlling thrips and the use of exclusion devices. Proceedings for the Eleventh Conference on Insect and Disease Management on Ornamentals, 18-20 February, 1995, Fort Meyers, FL. Society of American Florists, Alexandria, VA, pp. 113-120. Robb, K. L. 1998. Insecticide resistance management for ornamental plant protection. Proceedings for the Fourteenth Conference on Insect and Disease Management on Ornamentals, 21-23 February, 1998, Del Mar, CA. Society of American Florists, Alexandria, VA, pp. 67-75. Robb, K. L., C. Casey, A. Whitfield, and L. Campbell. 1998. A new weapon to fight INSV and TSVW. Grower Talks 61(12): 63-73. Robb, K. L., and M. P. Parrella. 1995. IPM of western flower thrips. In B. L. Parker, M. Skinner, and T. Lewis [eds.], Thrips Biology and Management. Plenum, New York, pp. 365-370. Ullman, D. E. 1996. Thrips and Tospoviruses: advances and future directions. Acta Horticulture 431: 310-324. Ullman, D. E. 1998. Managing Tospoviruses: developing a monitoring system for grower decision making. Proceedings for the Fourteenth Conference on Insect and Disease Management on Ornamentals, 21-23 February, 1998, Del Mar, CA. Society of American Florists, Alexandria, VA, pp. 9-12. Wawrzynski and Ascerno. In review. A survey of biological control use in Midwest greenhouse operations. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

![]()
Produced by Communication and Educational Technology Services, University of
Minnesota Extension Service.
In accordance with the Americans with Disabilities Act, this material is available in alternative formats upon request. Please contact your University of Minnesota Extension Service office or the Distribution Center at (800) 876-8636.
The University of Minnesota Extension Service is committed to the policy that all persons shall have equal access to its programs, facilities, and employment without regard to race, color, creed, religion, national origin, sex, age, marital status, disability, public assistance status, veteran status, or sexual orientation.